A new study in Open Biology explores the biological link between mood disorders and temperature rhythmicity using a mouse model of stress.

Problems with sleep and disturbances within circadian rhythms are commonly associated with various mental health disorders such as depression, anxiety, bipolar disorder, attention deficit hyperactivity disorder (ADHD), and bipolar disorder. A recent study by researchers at New York University Abu Dhabi investigates the effects of chronic social defeat stress (CSDS) to better understand the biological link between mood disorders and body temperature rhythmicity – lead author Dipesh Chaudhury tells us more.
Please could you tell us a little bit about your article?
There is a fair amount of anecdotal evidence that patients with mood disorders exhibit aberrations in diurnal rhythms in body temperature. However, the mechanisms by which stress and mood disorders lead to abnormal daily rhythms in body temperature is unknown. Understanding the cellular mechanisms connecting stress induced mood disorders with aberrations in daily rhythms in core body temperatures may open novel strategies for treating mood disorders. Moreover, there is a high comorbidity between metabolic disorders associated with obesity and mood disorders. Since metabolic processes are sensitive to changes in temperature, aberrations in daily rhythms in core body temperature may be the cause for disrupted metabolic processes leading to obesity. Thus, understanding these putative mechanisms may also lead to the treatment of obesity associated with mood disorders.
In our article we used a rodent model of stress, the CSDS paradigm to investigate the association between stress and daily rhythms in core body temperature. We found that both stress-resilient and -susceptible mice exhibited blunted rhythms during stress exposure and that body temperature of resilient mice recovers to baseline levels a little quicker then susceptible mice. We also investigated the effects of stress on expression of two thermosensitive genes, cold inducible RNA binding protein (CIRBP) and RNA-binding motif protein 3 (RBM3), in the lateral habenula (LHb), a brain region known to regulate mood. In control mice expression of CIRBP and RBM3 in the LHb is higher in the day when mice have lower body temperature compared to night when body temperature is higher. We found that immediately after stress exposure expression of CIRBP and RBM3 in the LHb were significantly blunted in both resilient and susceptible mice. Moreover, daily rhythmic expression of these 2 genes recovered to control levels 10-days after stress.
The preliminary findings of this study are novel and may help shed light on the interplay between temperature dysregulation, the clock system, and thermo-responsive gene expression in the LHb. Since CIRBP and RBM3 are able to effect expression of other target genes, it is possible that stress induced changes in molecular processes that lead to pathophysiological changes in brain circuits or metabolic pathways in the periphery may occur because of changes in rhythmic expression of these and other thermosensitive genes.
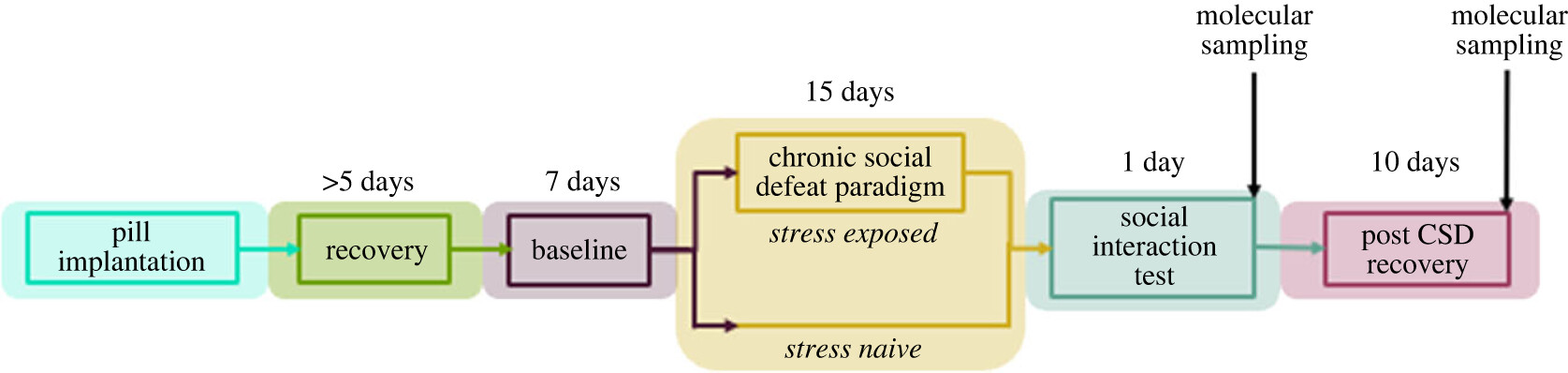
Timeline for investigating the association between stress, diurnal temperature rhythms and changes in expression of thermosensitive genes (Figure 1 doi.org/10.1098/rsob.220380)
What is the significance of the chronic social defeat stress (CSDS) model and why did you choose to use this?
The advantage of the CSDS paradigm is that we have two stress responsive phenotypes, where some mice are resilient to CSDS and others that are susceptible. The most common behavioural markers for resilient or susceptibility to CSDS is the social avoidance test. Young adult mice are social animals and when placed in an arena with other mice these young adults will typically want to interact with novel mice. Behaviourally, mice that are resilient to CSDS will continue to want to interact with novel mice after exposure to CSDS while mice susceptible to CSDS do not want to interact with novel mice. In other words, they usually isolate themselves and spend time alone. Over the last decades numerous published studies have reported physiologically and molecular differences between resilient and susceptible mice. The working model proposes that CSDS induces pathophysiological and molecular changes in brain circuits that leads to social isolation-like behaviours that loosely models aspects of mood disorders in susceptible mice. In contrast resilient mice are hypothesised to exhibit better homeostatic plasticity that buffers against stress induced pathophysiological and molecular changes observed in susceptible mice. Thus, by using the CSDS model not only can we investigate the effects of stress on physiological and molecular processes, we can also determine whether circadian processes such as core body temperature rhythms and sleep-wake cycle are differently affected in resilient and susceptible mice.
Were there any surprising findings from the study?
We had predicted that daily rhythms in core body temperature of resilient mice would likely recover faster than susceptible mice. This is probably indicative of better homeostatic plasticity in the resilient mice. We were however surprised that diurnal rhythms of the thermosensitive genes CIRBP and RBM3 recovered at the same rate in both resilient and susceptible mice 10-days after stress exposure. We had also measured Prok2 levels (this gene encodes a protein expressed in the suprachiasmatic nucleus (SCN) circadian clock) in the SCN in our study and found that its diurnal expression was not affected in resilient or susceptible mice because their levels were similar to control mice. Since Prok2 is an output molecule of the SCN that drive locomotor rhythms this would correlate with our observation that diurnal locomotor rhythms were not affected in resilient or susceptible mice. We were surprised that daily temperature, but not locomotor, rhythms were affected following exposure to CSDS.
What is next for your research, and where would you like to see research in the field heading?
This small paper is an initial part of our ongoing study into the effects of stress on daily rhythms in body temperature. We will continue to investigate the effects of stress on expression of CIRBP, RBM3 and other thermosensitive genes in the SCN, the region that that regulates circadian rhythms and other brain regions that regulate mood and sleep-wake cycle. We are also planning to investigate whether stress induced changes in expression of thermosensitive genes is associated with aberrations in metabolic processes linked to obesity.
We recently published a study showing differences in diurnal activity of lateral habenula cells projecting to the dorsal raphe nucleus where resilient mice exhibited robust daily rhythms in activity while susceptible mice exhibited significantly blunted rhythms. We are currently investigating whether diurnal differences in neural circuit in circuits connecting the SCN are responsible for these differences. We also recently published work that showed differences in aspects of the sleep-wake cycle between resilient and susceptible. The most exciting observation from these studies was evidence that mice that are susceptible to CSDS exhibit fragmented sleep prior to stress. This implies that fragmented sleep may be indicative of vulnerability to stress. Currently, we’re performing in-vivo recordings to determine whether differences in specific neural circuits that have a functional role in regulating sleep-wake transitions are responsible for the fragmented sleep observed in stress vulnerable mice.
Overall, I would like to see more detailed studies into the cellular and molecular mechanisms that drive stress induced changes in diurnal and circadian rhythms and sleep-wake cycle.
Do you have an exciting new discovery that you would like to publish in Open Biology? Find out more about our author benefits and submission process.
Image: Mouse covering its eyes with its paw, credit - sibya via Pixabay




